Team:Wageningen UR/Results
Results

On this page, we proudly present an birds-eye view of the results of the entire Xylencer project. Here you will find a summary of the most important conclusions drawn from each of the lines of research within Xylencer. In-depth reports on each topic can be found by clicking on their corresponding buttons below. For a more extensive report on our scientific highlights, including supporting figures, visit our demonstrate page.
Detection
The first pillar of Xylencer is focused on detecting X. fastidiosa by an automated tool enabling in-field testing based on Loop-mediated isothermal amplification (LAMP). Proof of principles for the key mechanisms of this device was obtained through multiple tests. We showed that we could perform LAMP on bacteria with X. fastidiosa specific primers. To ensure LAMP reagents stability over time and temperature influences, and to prevent insects from ingesting it, we showed that the reaction mixture can be preserved in paraffin at a slight reduction in short-term efficiency. By performing insect-transmission assays, we were able to show that insects fed on a plant mimic and could transfer bacteria from a source to this plant mimic. We determined the optimal feeding medium concentration, namely 0.2% sucrose, that still allows LAMP to function. Finally, in collaboration with The Netherlands food and consumer product safety authority (NVWA), we conducted a LAMP test on leaf extracts from infected plants. These tests demonstrated that a LAMP reaction temperature of 65 °C can lyse the bacterial cell wall and make their DNA accessible for the LAMP primers, successfully detecting X. fastidiosa with a visible readout.

Detection Device

Delivery
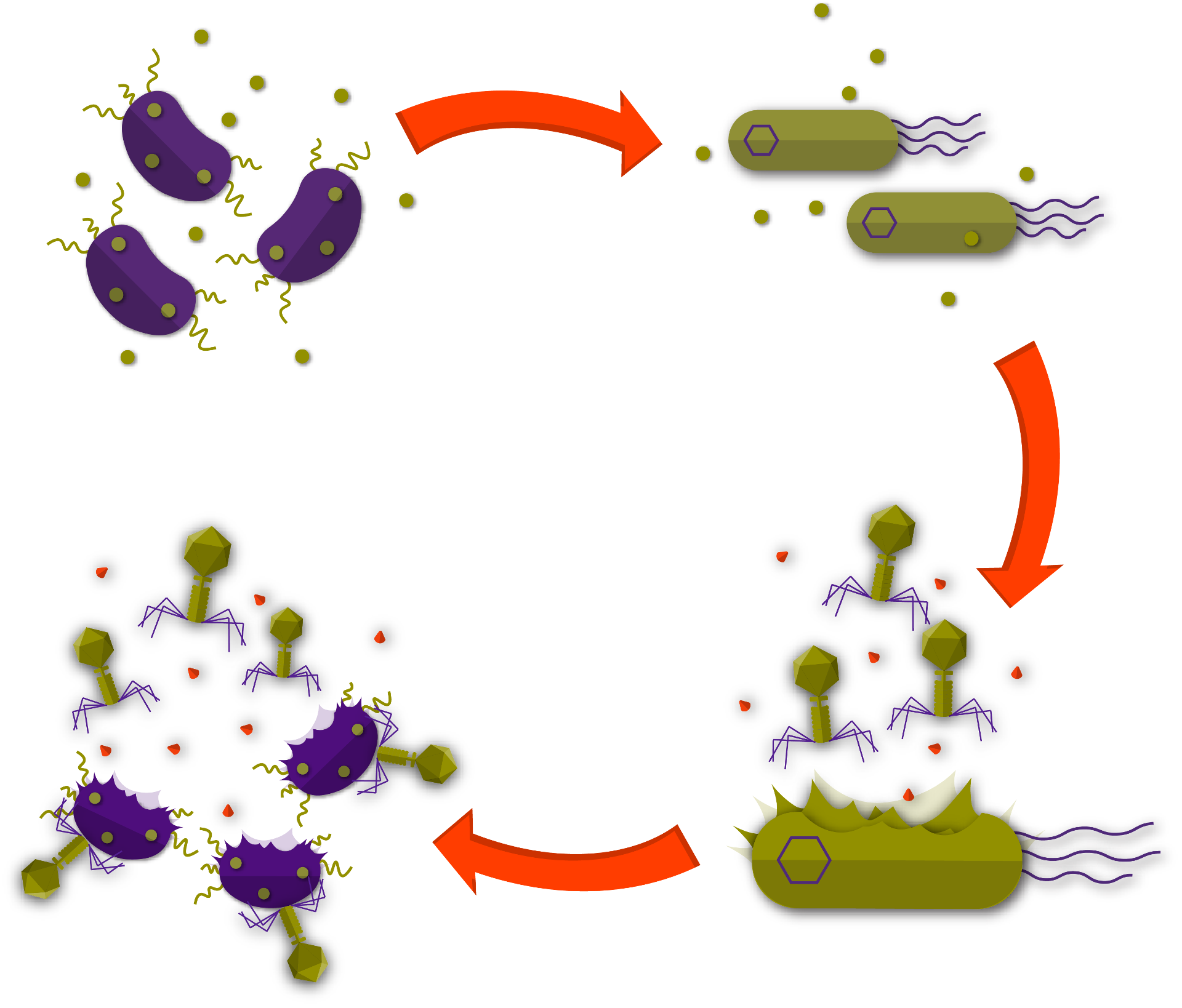
In the second pillar of Xylencer, we revolutionized phage delivery with the Phage Delivery Bacterium (PDB). To ensure our PDB would be safe, we trained a machine learning model on a manually curated pathogenicity dataset and established a genetic basis for non-pathogenicity. This allowed us to select Xanthomonas arboricola CITA 44 as our PDB. To direct the release of the pages to the areas of infection, we designed a circuit that controlled phage expression based on X. fastidiosa’s presence. For this circuit, we created a chimeric X. fastidiosa quorum-sensing receptor, linked to a tightly controlled riboswitch. This riboswitch is coupled to a CRISPR-dCas circuit that is designed to control phage expression. We showed that this system can control the expression of the phage operon. Besides this, we showed that dCas repression can be controlled with an Anti-CRISPR system, allowing us to fine-tune the circuit. To contain the spread of our PDB and guarantee biosafety, we explored the possibility of a timed kill-switch, that would only prove lethal after a set amount of time. We developed a circuit that increases protein expression over a period of three days. This system can be linked to a toxin/anti-toxin system to achieve delayed induced lethality.

Pathogenicity analysis

Sensing Circuit

Phage Repression

Kill Switch
Drag the Button to Reveal the PDB


Remediation
In the third pillar, remediation, we improved phage therapy by forging an alliance with the plant. This is achieved by alerting the plant to the presence of the pathogen, through artificially triggering a systemic plant immune response. To this end, Xylencer phages encode for a Pathogen Associated Molecular Pattern (PAMP): flg22. Phage infection and lysis of X. fastidiosa would set these PAMPs free within the plant’s xylem and trigger plant immune responses. We performed plant experiments by infecting Brassica oleacea with a close relative of X. fastidiosa, Xanthomonas campestris pv. Campestris and showed that inoculation of flg22 into the plant’s xylem decreases disease symptoms during the crucial early stages of infection.

Plant Immune Response

Spread
The final pillar includes the design of a self-spreading mechanism for the Xylencer phages. This allows our therapy to reach infected plants that would otherwise be missed by conventional methods. In order to achieve this, Xylencer phages will be bound to the mouthparts of the insects that spread X. fastidiosa by fusing adhesion proteins to their capsid. We demonstrated the ability of X. fastidiosa derived adhesion protein PD1764 to bind to structures found in the insect's mouthparts. Furthermore, we showed that we can produce a mix of wild-type and adhesion capsid fusion proteins, allowing for increased stability of the phage capsid by only partially saturating it with fusion proteins. Using cell-free transcription-translation we produced E. coli phage T7 and phage Lambda capsid proteins, serving as a model for the Xylencer phages. We constructed a spatial spread model that can simulate the effectivity and spread of the Xylencer phages. This model showed that our Xylencer phages would be effective at combatting X. fastidiosa, surpassing current methods.

Adhesion Library

Phage Engineering

Alternative Phage Engineering

Spatial Spread Model
