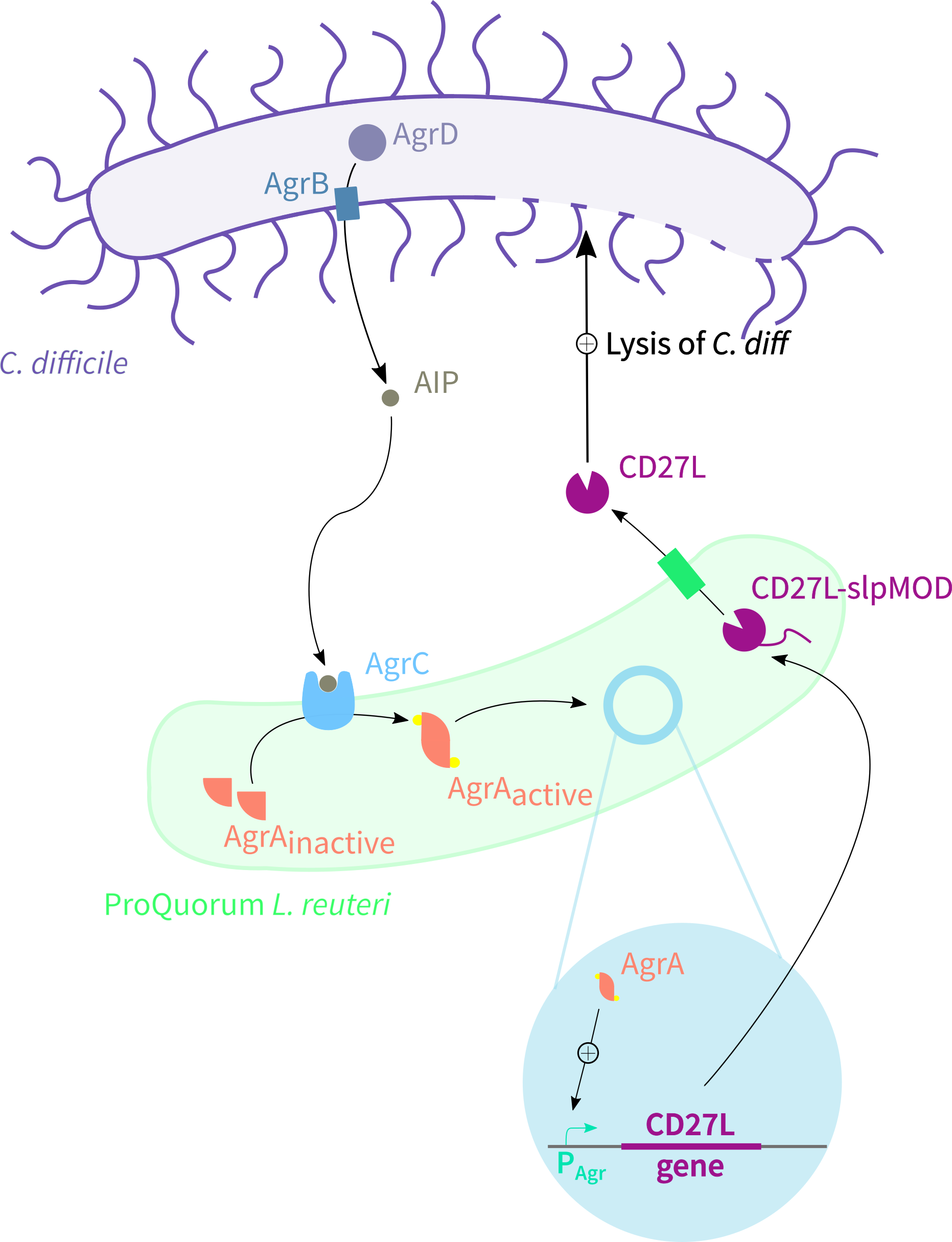
Team:Oxford/Model/Therapeutics-Insight

| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$M \to M + B$$ | $$K_b$$ | $$5.00 \times 10^{-14} \ hr^{-1}$$ |
| $$M \to M + D$$ | $$K_d$$ | $$5.00 \times 10^{-11} \ hr^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$B \to Bt$$ | $$\alpha_{Bt}$$ | $$3.334 \ hr^{-1}$$ | $$D \to Dt$$ | $$\alpha_{Dt}$$ | $$3.334 \ hr^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$Dt \overset{\mbox{Bt}}{\to} AIP$$ | $$k_S$$ | $$4.00 \times 10^8 \ M^{-1} hr^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$NULL \to M_e$$ | $$v$$ | $$1 \times 10^{2} \ hour^{-1}$$ |
| $$M_e \to M_e + E$$ | $$K_e$$ | $$4.65 \times 10^{-3} \ hour^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$X \to NULL \ (where X=B, D, Bt, Dt, S, me, E)$$ | $$\delta_X$$ | $$3.334 \ hr^{-1}$$ |
| Concentration Modelled | Differential Equation | Process Modelled |
|---|---|---|
| Cytosolic AgrB Protein | $$\frac {dB}{dt} = Kb \times M - (\alpha_{Bt} + \delta_{B}) \times B$$ | Translation + Degradation + Membrane Insertion |
| Cytosolic AgrD Protein | $$\frac {dD}{dt} = Kd \times M - (\alpha_{Dt} + \delta_{D}) \times D$$ | Translation + Degradation + Membrane Insertion |
| Transmembrane AgrB Protein | $$\frac {dB_t}{dt} = \alpha_B \times B - \delta_{Bt} \times Bt $$ | Membrane Insertion + Degradation |
| Transmembrane AgrD Protein | $$\frac {dD_t}{dt} = \alpha_Dt \times D - \delta_{Dt} - k_S \times Bt \times Dt$$ | Membrane Insertion + Degradation + Conversion into AIP |
| AIP | $$\frac {dS}{dt} =k_S \times Bt \times Dt + k_{ibind} \times R_{b_{cd}} - k_{bind} \times R_{cd} \times S + k_{ibind} \times R_{b_{lr}} - k_{bind} \times R_{lr} \times S - \delta_S \times S$$ | Conversion into AIP + Binding/Unbinding of AIP on C. difficile + Binding/Unbinding on L. reuteri + Binding + Degradation |
| Endolysin mRNA | $$\frac {dM_e}{dt} = v \times P - \delta_{M_e} \times M_e$$ | Transcription + Degradation |
| Endolysin Protein | $$Ke \times M_e - \delta_E \times E$$ | Translation + Degradation |
| Concentration Modelled | Differential Equation | Process Modelled |
|---|---|---|
| Population of L. reuteri | $$\frac {dN_l}{dt} = r_{lr} \times N_{t} \times (1- \frac {N_t}{N_{lr}})$$ | Logistic Growth of L. reuteri |
| Population of C. difficile | $$\frac {dN_c}{dt} = r_{cd} \times N_{c} \times (1- \frac {N_c}{N_{cd}})$$ | Logistic Growth of C. difficile |
| Total mRNA concentration in Population of L. reuteri | $$\frac {dM}{dt} = m \times N_{cd} - \delta_M \times M$$ | Transcription in Entire Population + Degradation |
| Total Endolysin concentration in Population of L. reuteri | $$\frac {dM_e}{dt} = v \times P \times N_{lr} - \delta_{M_e} \times M_e$$ | Transcription in Entire Population + Degradation |

Fig. 1 L. reuteri Growth Curve Assuming Initial Population = 1

Fig. 2 C. difficile Growth Curve Assuming Initial Population = 1

Fig. 3 Upregulation Profile of C. difficile and L. reuteri

Fig. 4 Growth Curves for Delayed C. difficile Growth

Fig. 5 Upregulation Profile of C. difficile and L. reuteri For Different Delay Times

















