Team:Oxford/Model/Design
| Endolysin Concentration Produced per L. reuteri cell /M CFU -1 |
|---|
| 0.00465 |
| Type of L. reuteri | Doubling time /hr | Growth Rate (ln 2/Doubling Time) /hr -1 |
|---|---|---|
| Wild Type | 0.548 | 1.26 |
| Consitutively Expressing CD27L Endolysin | 9.08 | 0.76 |

| Process Modelled | Differential Equation 1 | Explanation 1 | Differential Equation 2 | Explanation 2 |
|---|---|---|---|---|
| Logistic Growth Model of L. reuteri - Constitutive Expression | $$\frac {dC}{dt} = r_{stressed} \times C \times (1- \frac {C}{N_{lr}})$$ | Endolysin is being expressed constitutively. | N/A | N/A |
| Logistic Growth Model of L. reuteri - Dynamic Expression | $$\frac {dR_{1}}{dt} = r_{wildtype} \times R \times (1- \frac {R}{N_{lr}})$$ | If endolysin is not being expressed. | $$\frac {dR_{2}}{dt} = r_{stressed} \times R \times (1- \frac {R}{N_{lr}})$$ | If endolysin is being expressed. |
| Linear Expression of Endolysin | $$\frac {dEc}{dt} = k \times C$$ | For constitutive expression by L. reuteri | $$\frac {dEr}{dt} = k \times R$$ | For dynamic expression by L. reuteri |
| Colour of Graph Line | Explanation |
|---|---|
| Dotted | L. reuteri constitutively expressing endolysin. |
| Green | Endolysin yield increases with the time of induction of endolysin expression |
| Blue | Endolysin yield decreases with the time of induction of endolysin expression |
| Red | Endolysin yield of note is below endolysin yield from constitutive expression |
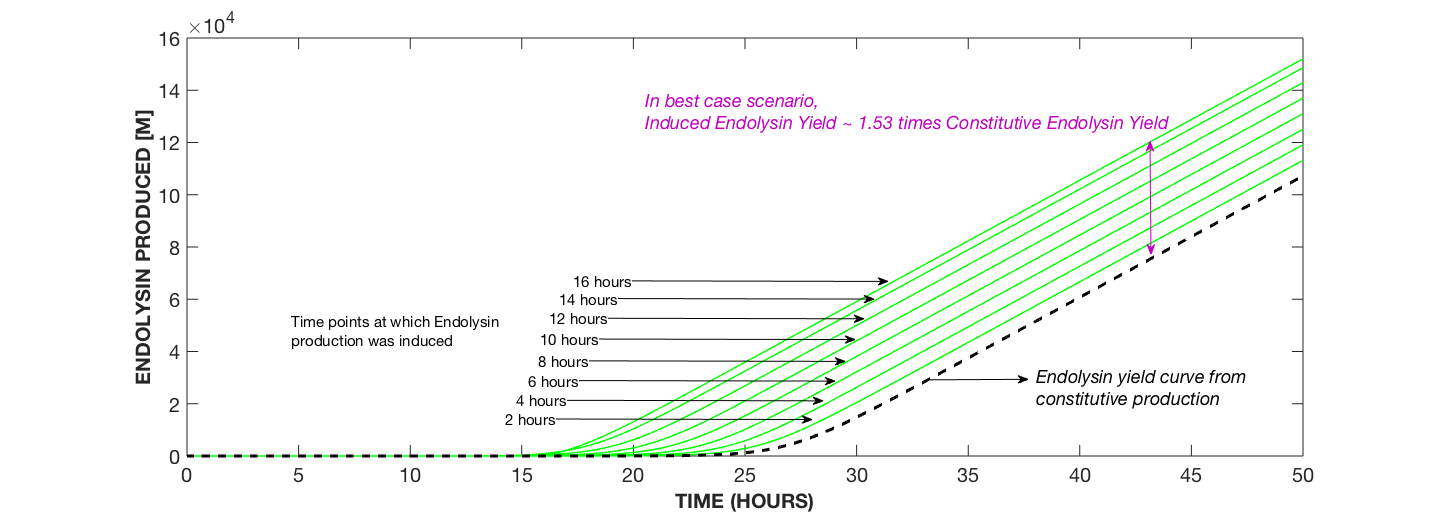
Fig. 1a compares the yield of endolysin from constitutive expression to that from dynamic expression whereby yield increases with increase in the time of induction.
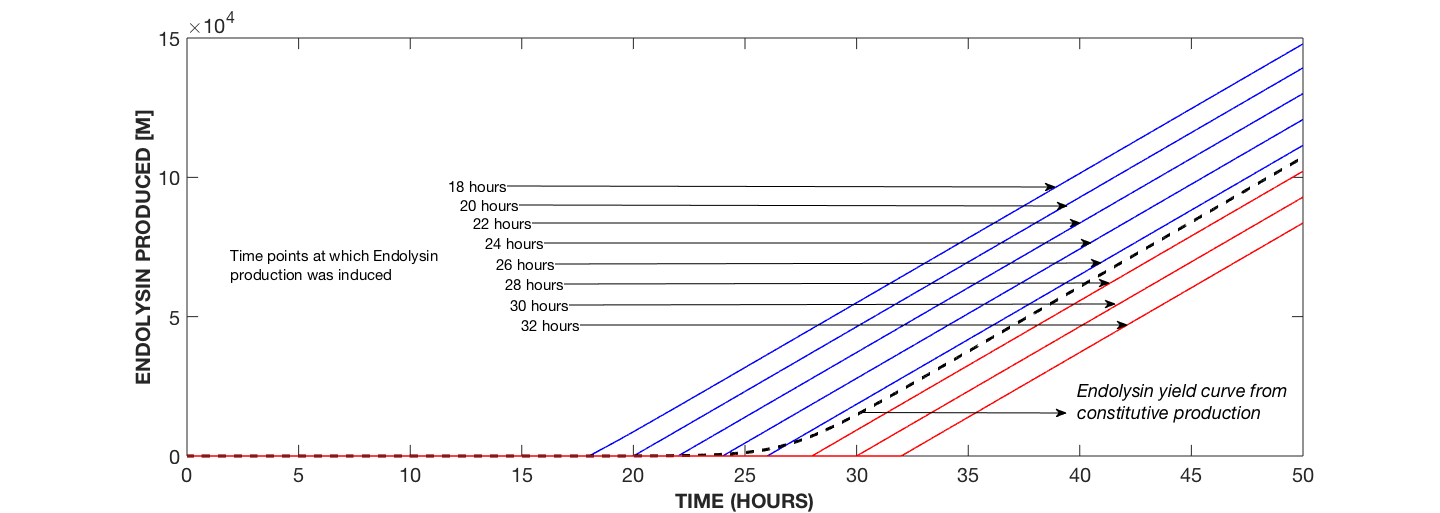
Fig. 1b compares the yield of endolysin from constitutive expression to that from dynamic expression whereby yield decreases with increase in the time of induction.
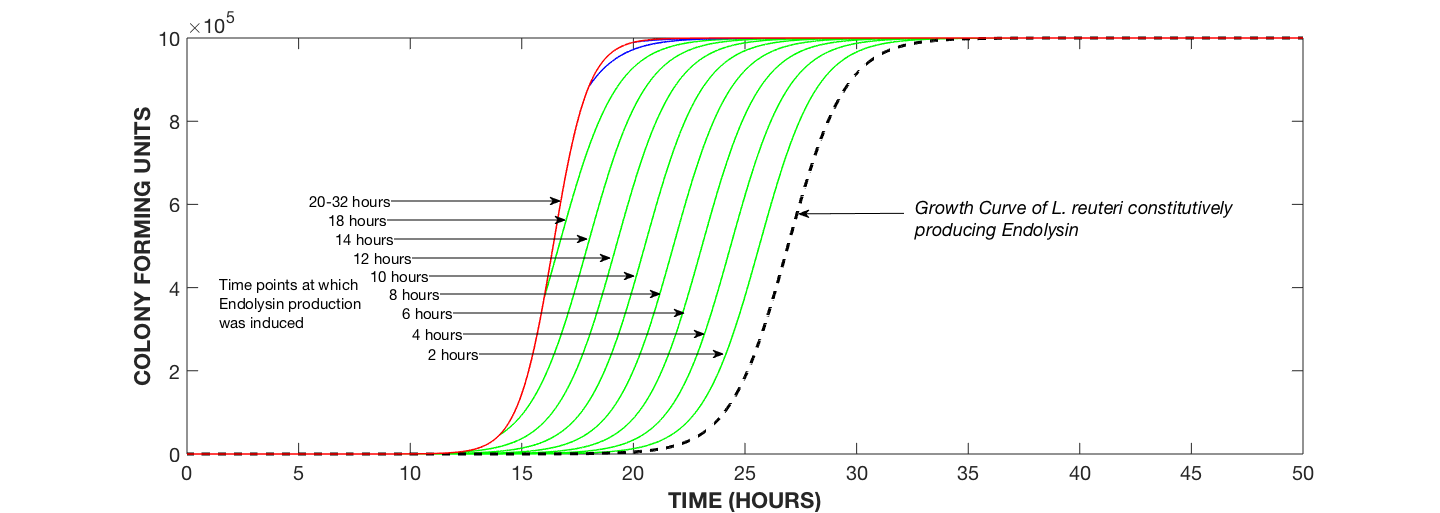
Fig 2: Growth Curves of L. reuteri Undergoing Endolysin Expression with Induction Times ranging from 2 hrs to 32 hrs.

| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$[NULL] \to M$$ | $$m$$ | $$8.33 \ M cell^{-1} \ minute^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | |
|---|---|---|---|
| $$M \to M + A$$ | $$K_a$$ | $$8.33 \times 10^{-8} \ minutes^{-1}$$ | |
| $$M \to M + C$$ | $$K_c$$ | $$8.33 \times 10^{-6} \ minutes^{-1}$$ | |
| $$C \to R$$ | $$\alpha_C$$ | $$0.0556 \ minutes^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | Inverse Kinetic Rate Constant | Inverse Kinetic Rate Constant Value Modelled |
|---|---|---|---|---|
| $$S + R \leftrightarrow R_b$$ | $$k_{bind}$$ | $$1.67 \times 10^{-7} \ (M \times minute)^{-1}$$ | $$k_{ibind}$$ | $$1.167 \ minutes^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | Inverse Kinetic Rate Constant | Inverse Kinetic Rate Constant Value Modelled |
|---|---|---|---|---|
| $$R_b \leftrightarrow R_{bp}$$ | $$k_{autop}$$ | $$6.3133 \ minutes^{-1}$$Ref. 3 | $$k_{iautop}$$ | $$ 10^{-2} \ minute^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | Inverse Kinetic Rate Constant | Inverse Kinetic Rate Constant Value Modelled |
|---|---|---|---|---|
| $$R_{b_p} + A \leftrightarrow R_b + A_p$$ | $$k_{phosph}$$ | $$5 \times 10^5 \ minutes^{-1}$$ | $$k_{dphosph}$$ | $$0.1777 \ minutes^{-1}$$Ref. 4 |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | Inverse Kinetic Rate Constant | Inverse Kinetic Rate Constant Value Modelled |
|---|---|---|---|---|
| $$2A_p \leftrightarrow A_{p_2}$$ | $$\phi_d$$ | $$3.33 \times 10^{8} \ (M \times minute)^{-1}$$ | $$\phi_m$$ | $$0.0167 \ minute^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$X \to NULL \ (where X=M, A, C, R, Rb, Rbp, S, Ap, Ap_2)$$ | $$\delta_X$$ | $$0.05566 \ minutes^{-1}$$Ref. 2 |
| Concentration Modelled | Differential Equation | Process Modelled |
|---|---|---|
| mRNA of AgrC Receptor & AgrA Protein | $$\frac {dM}{dt} = m - \delta_{M} \times M$$ | Transcription of AgrC & AgrA Genes + Degradation of mRNA |
| Deactivated AgrA Protein | $$\frac {dA}{dt} = K_a \times M - \delta_A \times A - k_{phosph} \times R_{bp} \times A + k_{dphosph} \times R_b \times A_p$$ | Translation of AgrA Protein + Degradation of AgrA Protein + Phosphorylation/Dephosphorylation of AgrA |
| Cytoplasmic AgrC Receptor | $$\frac {dC}{dt} = K_a \times M - \alpha_C \times C - \delta_C \times C$$ | Translation of AgrC Receptor + Membrane Insertion of AgrC Receptor + Degradation of AgrC Receptor |
| Unbound AgrC Receptor | $$\frac {dR}{dt} = \alpha_C \times C - \delta_R \times R + k_{ibind} \times R_b - k_{bind} \times R \times S$$ | Membrane Insertion of AgrC Receptor + Degradation of Inserted AgrC Receptor + Binding/Unbinding of AgrC Receptor |
| Bound AgrC Receptor | $$\frac {dR_b}{dt} = k_{bind} \times S \times R - k_{ibind} \times R_b - \delta_{R_b} \times R_b + k_{iauto} \times R_{bp} - k_{autop} \times R_b + k_{phosph} \times R_{b_p} \times A - k_{dphosph} \times R_b \times A_p$$ | Ligand Binding/Unbinding of AgrC Receptor + Degradation of Bound AgrC Receptor + Autophosphorylation/De-autophosphorylation + Phosphotransfer/Dephosphorylation of AgrC Receptor |
| Phosphorylated AgrC Receptor | $$\frac {dR_{b_p}}{dt} = k_{autop} \times R_b - k_{iauto} \times R_{b_p} - k_{phosph} \times R_{b_p} \times A + k_{dphosph} \times R_b \times A_p - \delta_{Rb_p} \times R_{b_p}$$ | Autophoshorylation/Auto-dephosphorylation of AgrC Receptor + Phosphotransfer/Dephosphorylation to AgrA Protein + Degradation of Phosphorylated AgrC Receptor |
| AgrA Protein | $$\frac {dA_p}{dt} = k_{phosph} \times R_{bp} \times A - k_{dphosph} \times R_b \times A_p - 2 \times \phi_d \times A_{p}^2 + \phi_m \times A_{p_2} - \delta_{A_p} \times A_p$$ | Phosphotransfer/Dephosphorylation of AgrA Protein + Dimerisation/Monomerisation + Degradation of AgrA Protein |
| Dimerised AgrA Protein | $$\frac {dA_{p_2}}{dt} = \phi_d \times A_{p}^2 - 0.5 \times \phi_m \times A_{p_2} - \delta_{Ap_2} \times A_{p_2}$$ | Dimerisation/Monomerisation of AgrA Protein + Degradation of Dimerised AgrA Protein |
| Parameter | Explanation |
|---|---|
| b | Binding Rate of Transcription Factor |
| u | Unbinding Rate of Transcription Factor |
| Ap | Population-wide Concentration of Transcription Factor in the S. aureus Quorum Sensing System |
| N | Number of Cells |
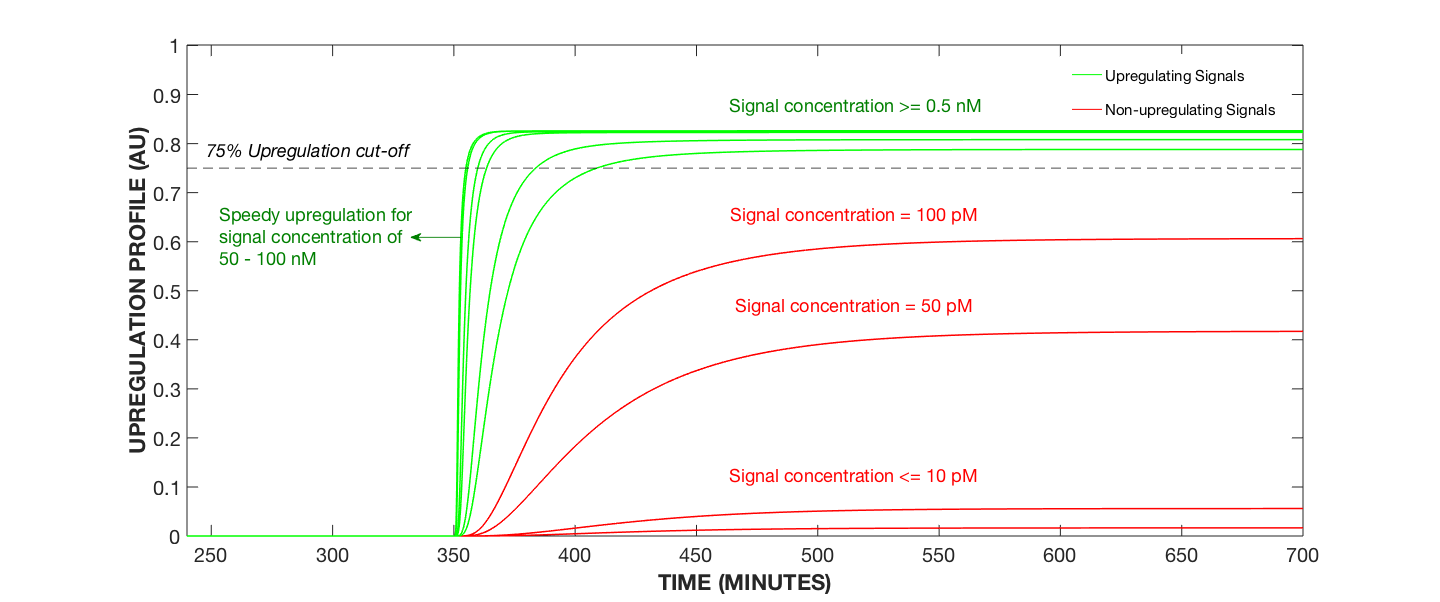
Figure 1. Upregulation Profile for Varying AIP Concentrations
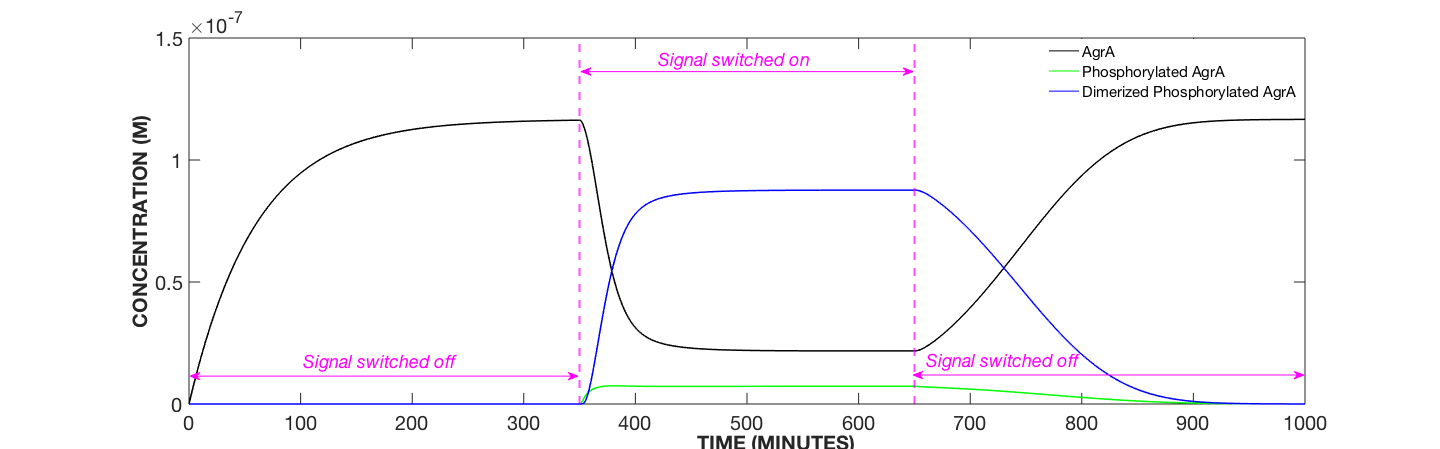
Figure 2. Transcription Factor Dynamics, AIP Concentration Modulated
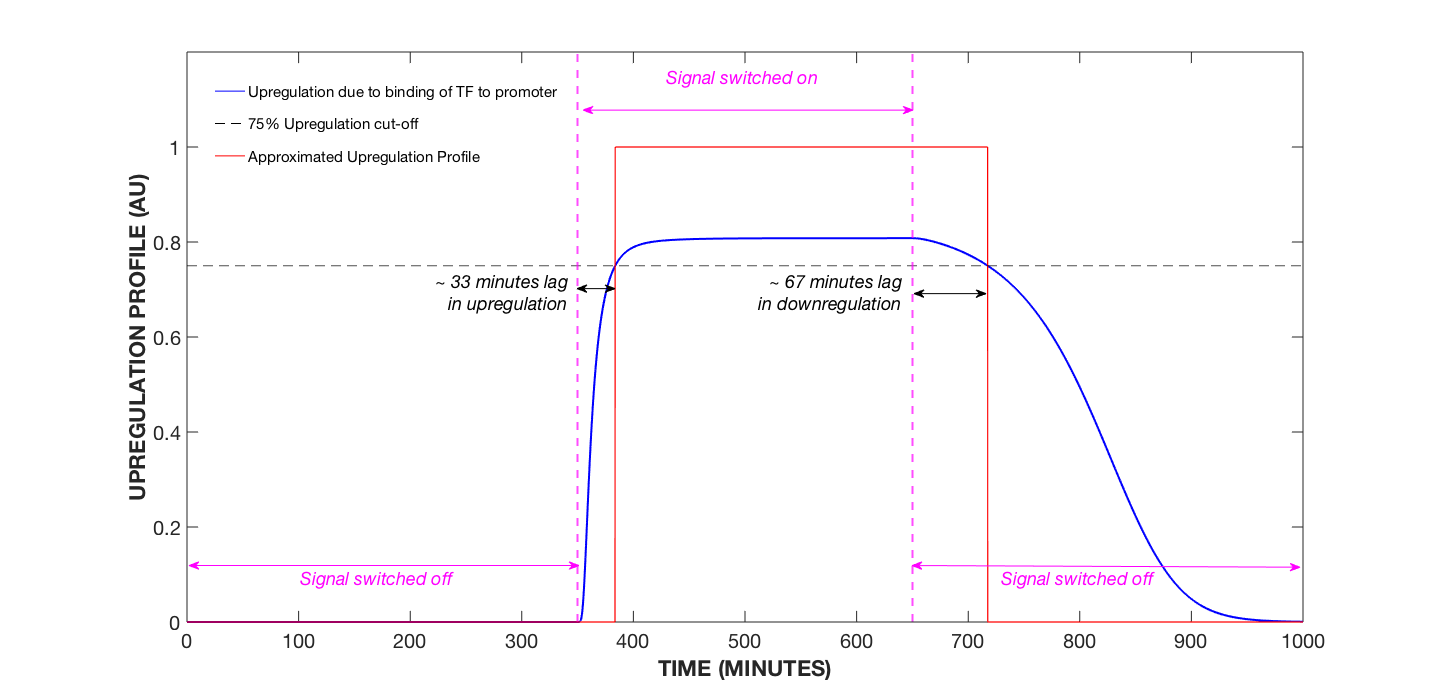
Figure 3. Upregulation Profile of Cell, AIP Concentration Modulated
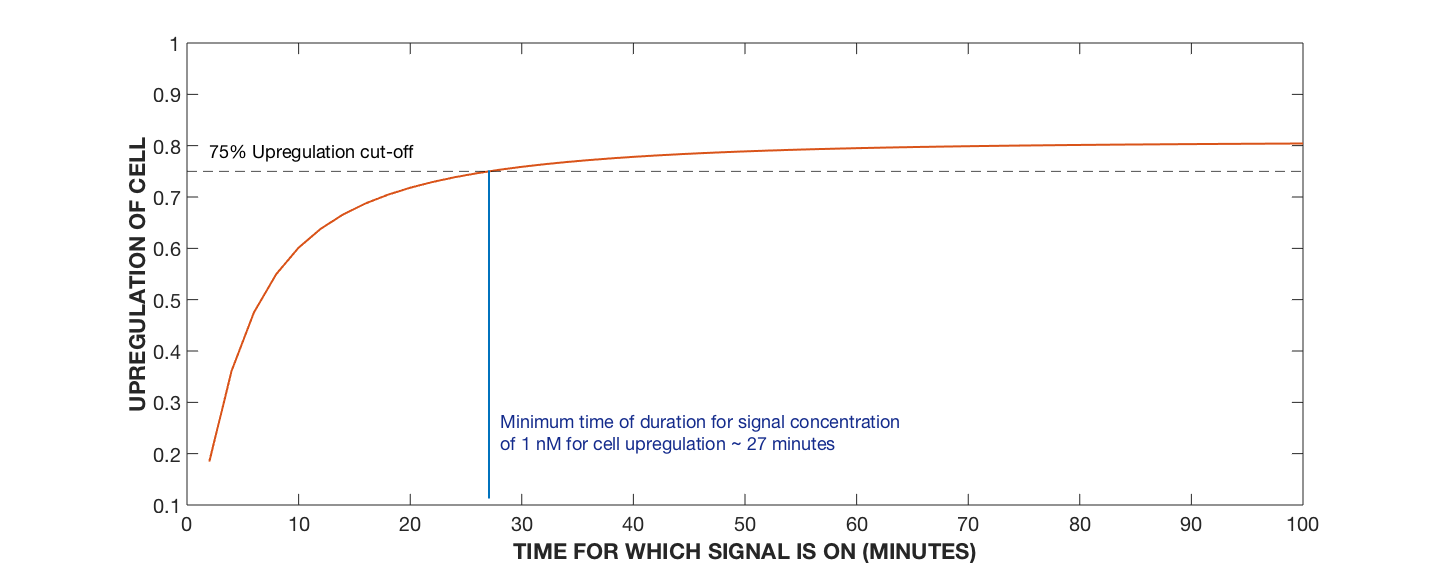
Figure 4. Upregulation Profile of Cell, Varying AIP Signalling Time
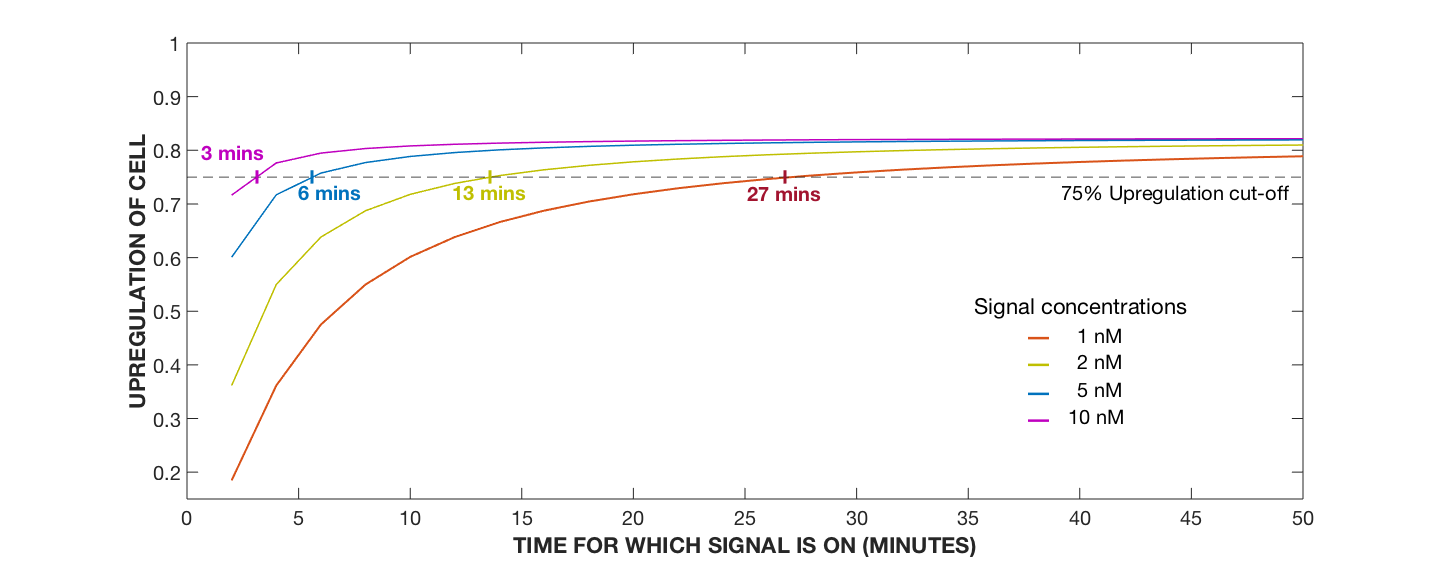
Figure 5. Upregulation Profile of Cell, Varying AIP Signalling Time with Varying AIP Concentrations
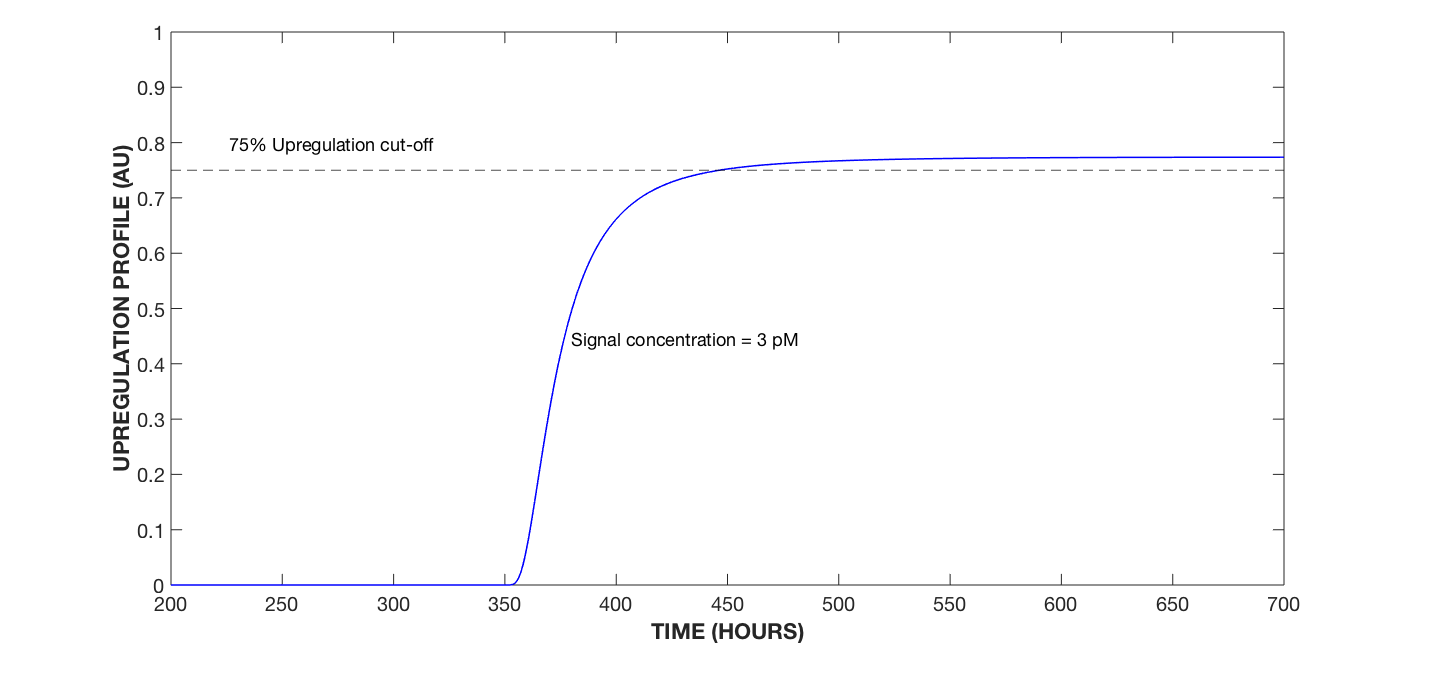
Figure 6. Upregulation Profile of Cell, 3 pM AIP Signal Concentration
| # | Reference |
|---|---|
| 1 | Jabbari, Sara, et al. “Mathematical Modelling of the Agr Operon in Staphylococcus Aureus.” Journal of Mathematical Biology, vol. 61, no. 1, 2009, pp. 17–54., doi:10.1007/s00285-009-0291-6. |
| 2 | Wright, J. S., et al. “Transient Interference with Staphylococcal Quorum Sensing Blocks Abscess Formation.” Proceedings of the National Academy of Sciences, vol. 102, no. 5, 2005, pp. 1691–1696., doi:10.1073/pnas.0407661102. |
| 3 | Srivastava, S. K., et al. “Influence of the AgrC-AgrA Complex on the Response Time of Staphylococcus Aureus Quorum Sensing.” Journal of Bacteriology, vol. 196, no. 15, 2014, pp. 2876–2888., doi:10.1128/jb.01530-14. |
| 4 | Wang, Boyuan, and Tom W. Muir. “Regulation of Virulence in Staphylococcus Aureus : Molecular Mechanisms and Remaining Puzzles.” Cell Chemical Biology, vol. 23, no. 2, 2016, pp. 214–224., doi:10.1016/j.chembiol.2016.01.004. |

















