Team:Oxford/Model/Further-Development
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$C. difficile_{downregulated} \to C. difficile_{downregulated} + AIP$$ | $$k_S$$ | $$10^{3} \ M \ S^{-1}$$ |
| $$AIP \to [NULL]$$ | $$\delta_AIP$$ | $$10^{2} \ S^{-1}$$ |
| $$C. difficile_{downregulated} + AIP \to C. difficile_{upregulated}$$ | $$\phi_c$$ | $$10^{6} \ M^{-1} \ S{-1}$$ |
| $$L. reuteri_{downregulated} + AIP \to L. reuteri_{upregulated}$$ | $$\phi_l$$ | $$10^{6} \ M^{-1} \ S{-1}$$ |
| Species | Diffusion Rate Constant Value Modelled |
|---|---|
| $$AIP$$ | $$10^{-9} \ cm^{2} \ s^{-1}$$ |
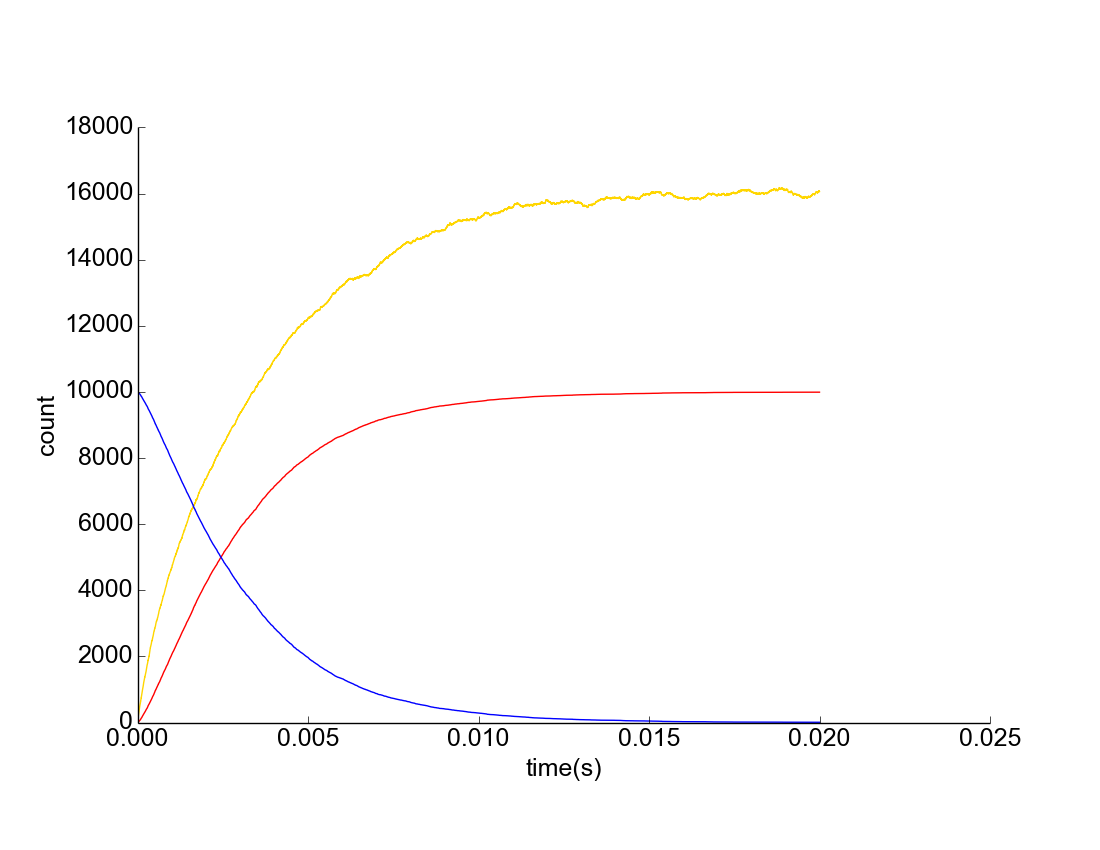
Figure 1. Upregulation of C. difficile Assuming Uniform Mixture. Yellow represents AIP concentration, Blue represents downregulated C. difficile population, Red represents upregulated C. difficile population.
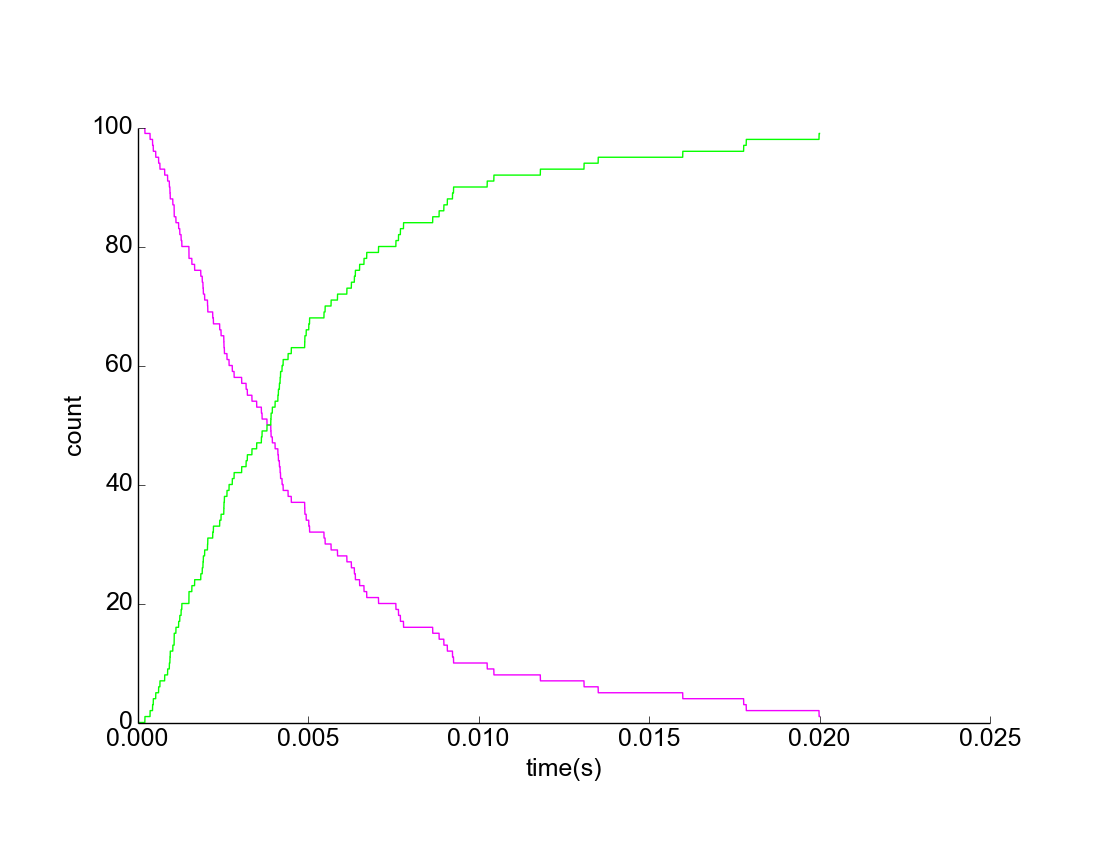
Figure 2. Upregulation of L. reuteri Assuming Uniform Mixture. Purple represents downregulated L. reuteri population; Green represents upregulated L. reuteri population.
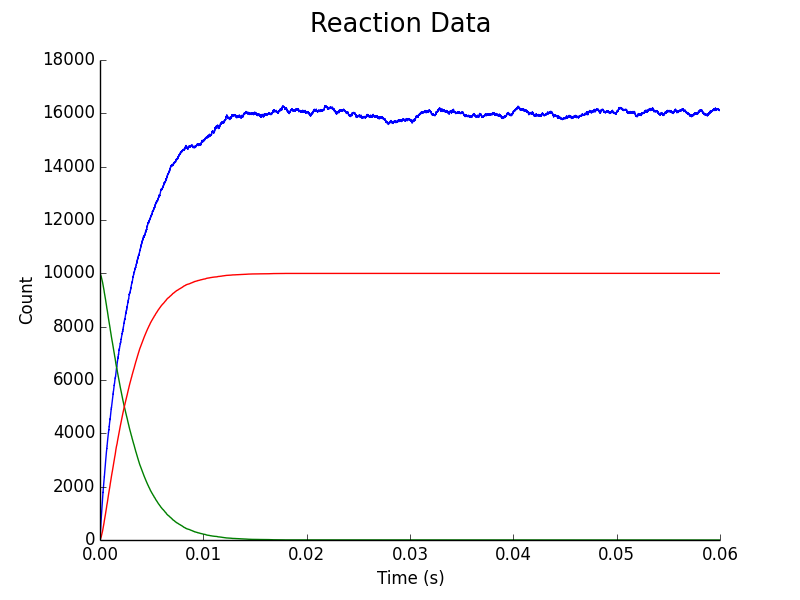
Figure 3. Upregulation of C. difficile assuming C. difficile & L. reuteri Colonies Adjacent. Blue represents AIP concentration, Green represents downregulated C. difficile population, Red represents upregulated C. difficile population.
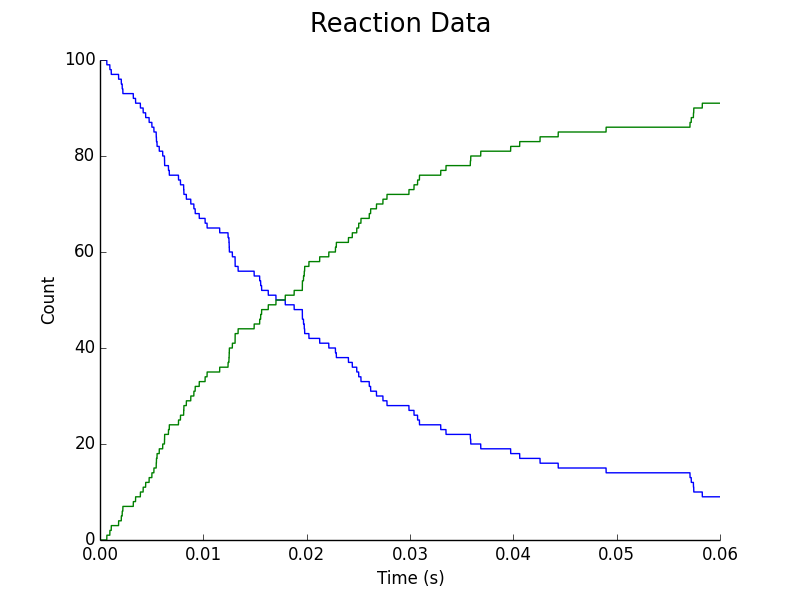
Figure 4. Upregulation of L. reuteri assuming C. difficile & L. reuteri Colonies Adjacent. Blue represents downregulated L. reuteri population; Green represents upregulated L. reuteri population.
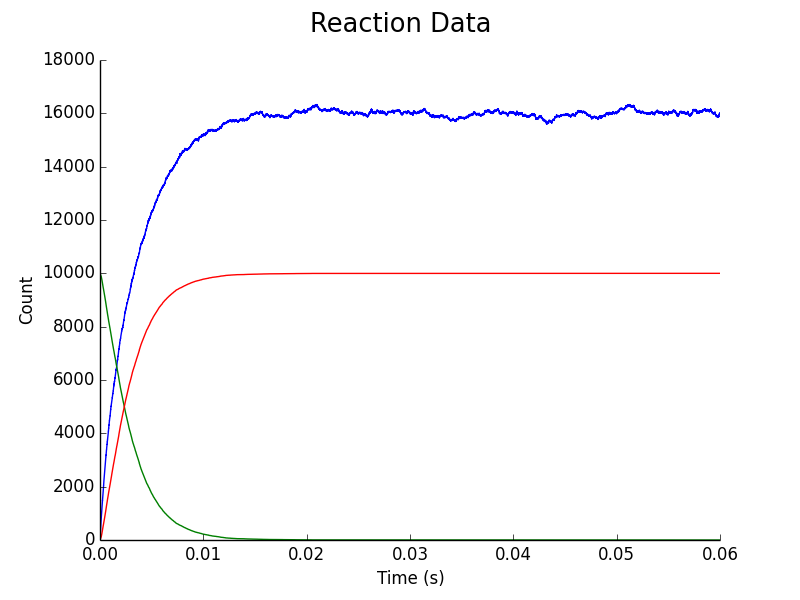
Figure 5. Upregulation of C. difficile assuming C. difficile & L. reuteri colonies are separated by a fixed distance in space. Blue represents AIP concentration; Green represents downregulated C. difficile population; Red represents upregulated C. difficile population.
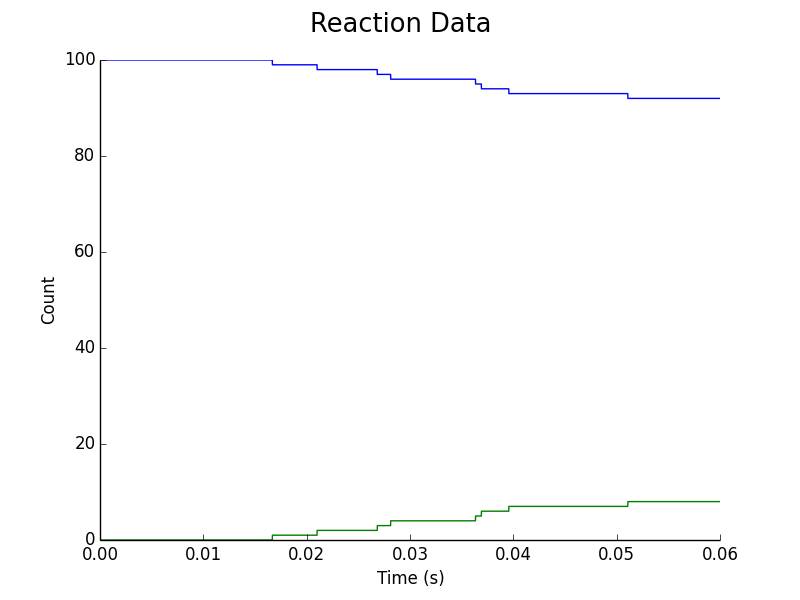
Figure 6. Upregulation of L. reuteri assuming C. difficile & L. reuteri colonies are separated by a fixed distance in space. Blue represents downregulated L. reuteri population, Green represents upregulated L. reuteri population.
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$L. reuteri_{downregulated} \to L. reuteri_{downregulated} + S$$ | $$k_S$$ | $$10^{3} \ M \ s^{-1}$$ |
| $$S \to [NULL]$$ | $$\delta_S$$ | $$10^{2} \ s^{-1}$$ |
| $$L. reuteri_{downregulated} + S \to L. reuteri_{upregulated}$$ | $$\gamma$$ | $$10^{6} \ M^{-1} \ s^{-1}$$ |
| Species | Diffusion Rate Constant Value Modelled |
|---|---|
| $$AIP$$ | $$10^{-9} \ cm^{2} \ s^{-1}$$ |
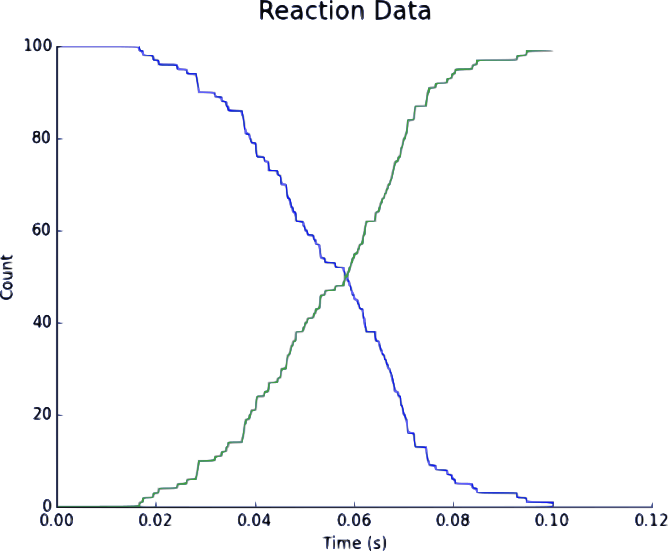
Figure 1. Upregulation of L. reuteriwith a Secondary Signalling System when C. difficile & L. reuteri are separated by a fixed distance in space. Blue represents downregulated L. reuteri population; Green represents upregulated L. reuteri population.

| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$NULL \to M_{xy}$$ | $$m_2 \ (if [Ap_2] \geq 6.514 \times 10^{-9} \ M); \\ 0 \ (if [Ap_2] \lt 6.514 \times 10^{-9} \ M)$$ | $$8.33 \ M \ hour^{-1} \ cell^-1$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$M_{xy} \to X$$ | $$v$$ | $$1.67 \times 10^{-7} \ minutes^{-1}$$ |
| $$M_{xy} \to Y$$ | $$v$$ | $$1.67 \times 10^{-7} \ minutes^{-1}$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$X + Y \to XY$$ | $$k_{ir}$$ | $$0.0167 \ M^{-1} \ minutes^{-1}$$ |
| Kinetic Rate Constant | Kinetic Rate Constant Value Modelled | |
|---|---|---|
| $$P$$ | $$1 \ (if XY \geq 0.25 \times 10^{-5} \ M) \ or \ 0 \ (if XY \lt 0.25 \times 10^{-5} \ M)$$ |
| Kinetic Rate Equation | Kinetic Rate Constant | Kinetic Rate Constant Value Modelled |
|---|---|---|
| $$X \to NULL \ (where X=M_{xy}, X, Y)$$ | $$\delta_X$$ | $$0.0556 \ hour^{-1}$$ |
| Concentration Modelled | Differential Equation | Process Modelled |
|---|---|---|
| mRNA for Signaling Molecule X & Receptor Y | $$\frac {dM_{xy}}{dt} = m_2 - \delta_{M_{xy}} \times M_{xy}$$ | Transcription + Degradation |
| Signalling Molecule X | $$\frac {dX}{dt} = v \times M_{xy} - k_{ir} \times X \times Y - \delta_X \times X$$ | Translation + Ligand Binding + Degradation |
| Signalling Molecule Y | $$\frac {dY}{dt} = v \times M_{xy} - k_{ir} \times X \times Y - \delta_Y \times Y$$ | Translation + Ligand Binding + Degradation |
| Ligand-Receptor Complex XY | $$\frac {dXY}{dt} = k_{ir} \times X \times Y - \delta_{XY} \times XY$$ | Ligand Binding + Degradation |
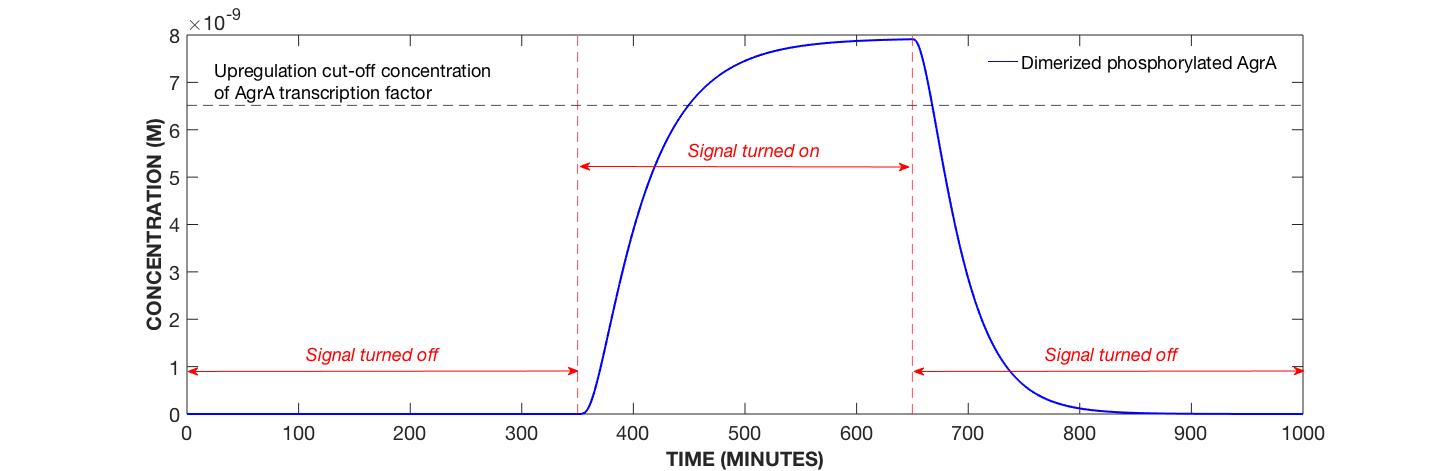
Figure 1. Dynamics of Dimerised, Phosphorylated AgrA Protein with Secondary Amplification System, AIP Concentration Modulated. Specifically, AIP added at 350 minutes and removed at 650 minutes.
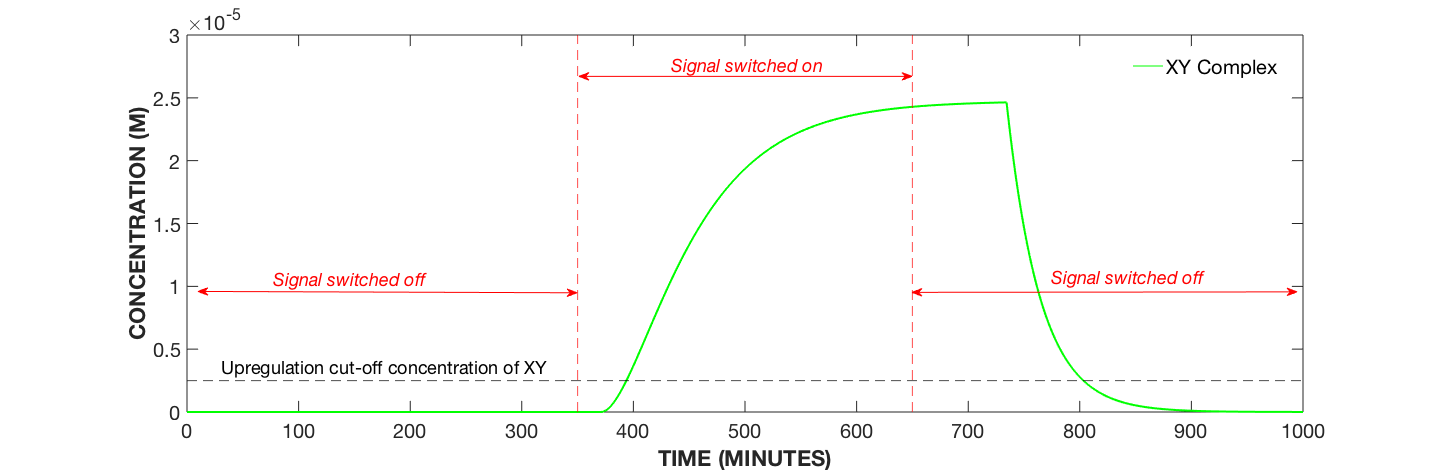
Figure 2. Dynamics of Signal-Receptor Complex XY with Secondary Amplification System, AIP Concentration Modulated. Specifically, AIP added at 350 minutes and removed at 650 minutes.
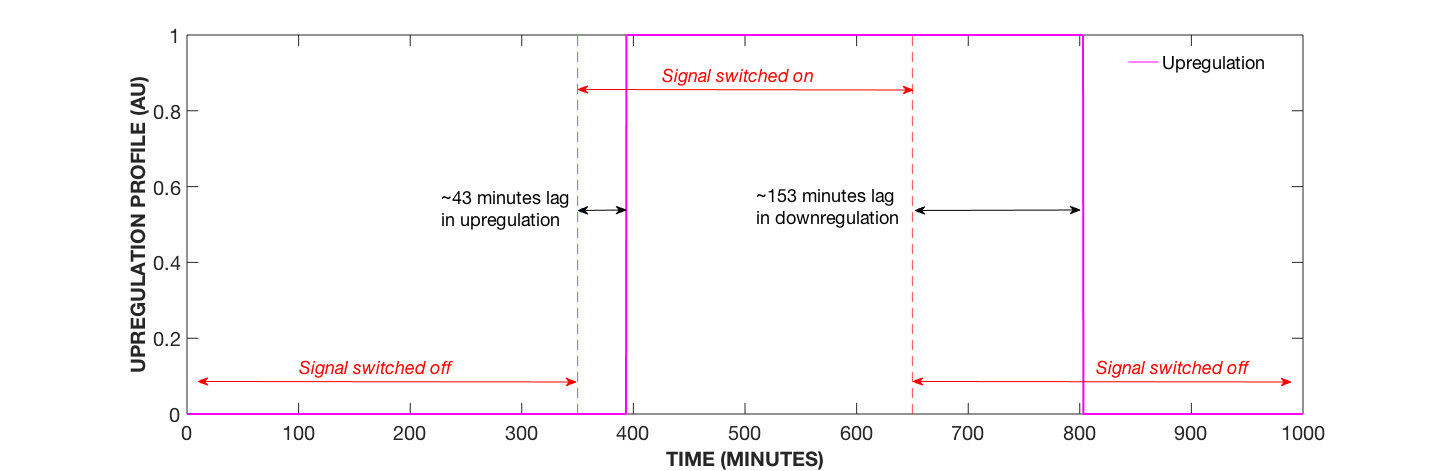
Figure 3. Upregulation Profile with Secondary Amplification System, AIP Concentration Modulated. Specifically, AIP added at 350 minutes and removed at 650 minutes.
| Reference |
|---|
| Stiles, JR, et al. (1996). Miniature endplate current rise times < 100 μs from improved dual recordings can be modeled with passive acetylcholine diffusion from a synaptic vesicle. Proc. Natl. Acad. Sci. USA 93:5747-5752. |
| Jabbari, Sara, et al. “Mathematical Modelling of the Agr Operon in Staphylococcus Aureus.” Journal of Mathematical Biology, vol. 61, no. 1, 2009, pp. 17–54., doi:10.1007/s00285-009-0291-6. |
| Stiles, JR, and Bartol, TM. (2001). Monte Carlo methods for simulating realistic synaptic microphysiology using MCell. In: Computational Neuroscience: Realistic Modeling for Experimentalists, ed. De Schutter, E. CRC Press, Boca Raton, pp. 87-127. |
| Kerr R, Bartol TM, Kaminsky B, Dittrich M, Chang JCJ, Baden S, Sejnowski TJ, Stiles JR. (2009). Fast Monte Carlo Simulation Methods for Biological Reaction-Diffusion Systems in Solution and on Surfaces. SIAM J. Sci. Comput., 30(6):3126-3149. |

















