This Page Contains (Structural Modeling and T-cell Kinetics Modeling)
A- Structural Modeling
· Computational Antibody Design
1- A library of commercial and experimental antibodies have been retrieved form IMGT [1] database and was modeled using oxford antibody modelling tool [2] to form a library of 3D structures of scFv(s) of retrieved antibodies.
2- Amino acid sequences and structures of estrogen receptor beta (ESR2) and Major Egg antigen (MEA) of Schistosoma haematobium were retrieved from uniprot database and s haematobium database and remodeled using Robetta [3].
3- As ESR2 is an Intracellular antigens is non-exclusively expressed in bladder cancer cells, thus we had to design our antibodies to target a specific variant of ESR2. To achieve this we performed a neoantigen screening through can neoantigen database [4] to select specific bladder cancer variant of ESR2 (variant R329Q) for HLA-A*02:01.
4- The robetta generated models of ESR2 (including mutated structure bladder cancer specific variant R329Q) and Major Egg antigens were submitted to IEDB MHC-I epitope prediction recommended method [5] to select most stable presented epitope against HLA-A*02:01 to targeted as ESR2 is an intracellular antigen.
5- The generated library of scFv(s) were docked against a modelled structures of both ESR2 and MEA using PatchDock global docking tool [6] as an initial screening step for selecting potential antibodies.
6- The top 10 generated complexes of scFv with both antigens ESR2 and MEA were selected and subjected to specific local docking using Rosetta antibody docking [7].
7- Then, the most efficient structure was subjected to alanine scanning, to further specify the residues responsible for binding using Rosetta alanine scanning [8] , The top antibody according to scores and binding affinity was further subjected to generation of random mutation [9] libraries according to conservation matrices of residues of each heavy and light chains in top scFv for both targets.
8- The generated libraries were further redocked using Rosetta docking to select the most stable mutated structure.
9- Molecular Dynamic simulations MDs were used through Amber99sb-ildn force field and TIP3P water model to judge the stability of the model regarding both light and heavy chains interactions [10].
10- The selected 2 antibodies were subjected to codon optimization on benching, to optimize it for DNA synthesis and further construction of chimeric antigen receptor CAR construct.
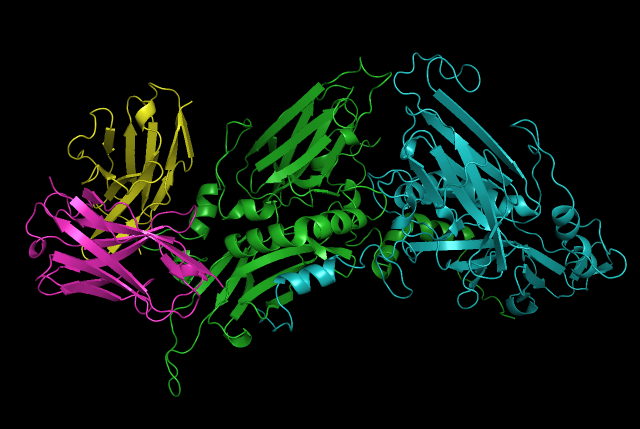
ESR2 Designed Antibody ScFv in binding pose with ESR2 (H,L chains in Purple and Yellow respectively, while estrogen-receptor beta chains are in green and cyan)
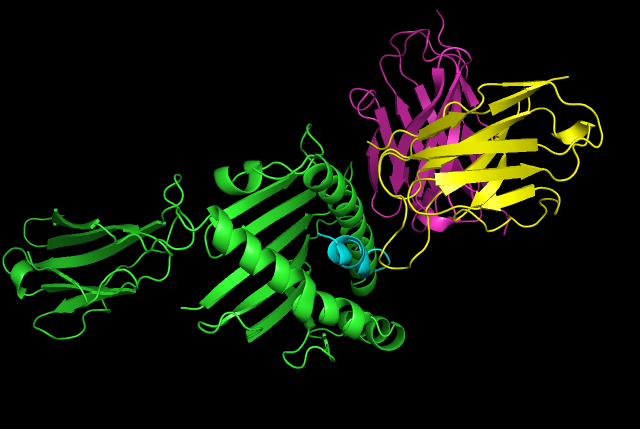
Major Egg antigen of s.haematobium ova Designed Antibody ScFv in binding pose with ESR2 (H,L chains in Purple and Yellow respectively, while MEA chains are in green and cyan(antigenic fragment))
· Structural Modelling of Pharmacological switch-off
Safety is one of the main issues that hinders the applicability of T-cell based therapies, due to expected undesired hyper-activation of Immune response as clinically observed in Cytokine Release syndromes with CAR T-cell treated patients. Thus, it’s critical to design an efficient switch off mechanism for the T-cell system in case of undesired storm of cytokines that could be fatal in clinical application.
Mestermann et al. found that dasatinib –a tyrosine kinase inhibitor- potently inhibited the autophosphorylation of LCK, a kinase that phosphorylates the T cell receptor (TCR) CD3 ζ-chain. Accordingly, activatory phosphorylation of the CAR CD3ζ (zeta) domain and the CD3ζ-associated kinase ZAP70 was reduced by ~90%, which prevented downstream NFAT-driven gene expression. Dasatanib was equally effective as an off switch regardless of the CAR target antigen (CD19, ROR1 or SLAMF7) or costimulatory domain (CD28 or 4-1BB). The drug was less effective, however, if added >1 hour after CAR T cell stimulation, but the results of sequential stimulation experiments suggested that most CAR T cells would eventually be inhibited upon subsequent encounters with target cells. Importantly, CAR T cells regained their full anticancer activity rapidly after dasatinib withdrawal, even after a week of exposure [11].

In our Modeling section, we investigated the interaction between dasatinib and LCK in order to reveal it’s potential as a safe pharmacological inhibitor of clinical cytokine release complications of CAR-T cell therapy. which revealed potential -8.3 Kcal/mol interaction between dasatinib and key residues of phosphorylation of lck including Tyr394 with vina-based blind docking algorithm cb-dock [12].
· B- CAR Kinetics Modeling
This model has been based on non-linear ODE (ordinary differential equations) through a predator-prey bio mathematical model of CAR T-cells killing activity against cancer cells.
Let X = Cancer cell density
Y = CAR T-cells density
Z = Il-18 Expression level
Z= TRUCK T-Cells (Il-18 expressing CAR T-cells)
Following this set of non-linear ODEs

Table of Constant parameters (Patrivia et al [13])
Parameter | Description | Unit |
ρ | Cancer cell net growth rate | day-1 |
Κ1 | CAR T-cell killing rate | day-1 cell-1 |
Κ2 | Net rate of proliferation and exhaustion of T-cells | day-1 cell-1 |
θ | CAR T-cell net death | day-1 |
Κz | Il-18 TRUCK killing rate | day-1 cell-1 |
In order to inspect the intrinsic dynamics of the system we scaled the X and Y cellular populations as well as time to remove physical units, so that steady-state could be obtained by setting time derivative to zero in equilibrium.

The model assumes that Cellular populations are well mixed, where cancer cell grows logistically while CAR-T cells grow independently of cancer cells and interaction between CAR T-cells and cancer cells follows the law of mass action and does not follow variations in antigen density. Proliferation of CAR T-cells could also define T-cells exhaustion.
What is the Optimum Effector to Target E:T ratio?
We have generated different runs to test each E:T ratio and we found that killing rate decreases with increasing the E:T ratio and T-cell doses which is directly correlated to CAR exhaustion.
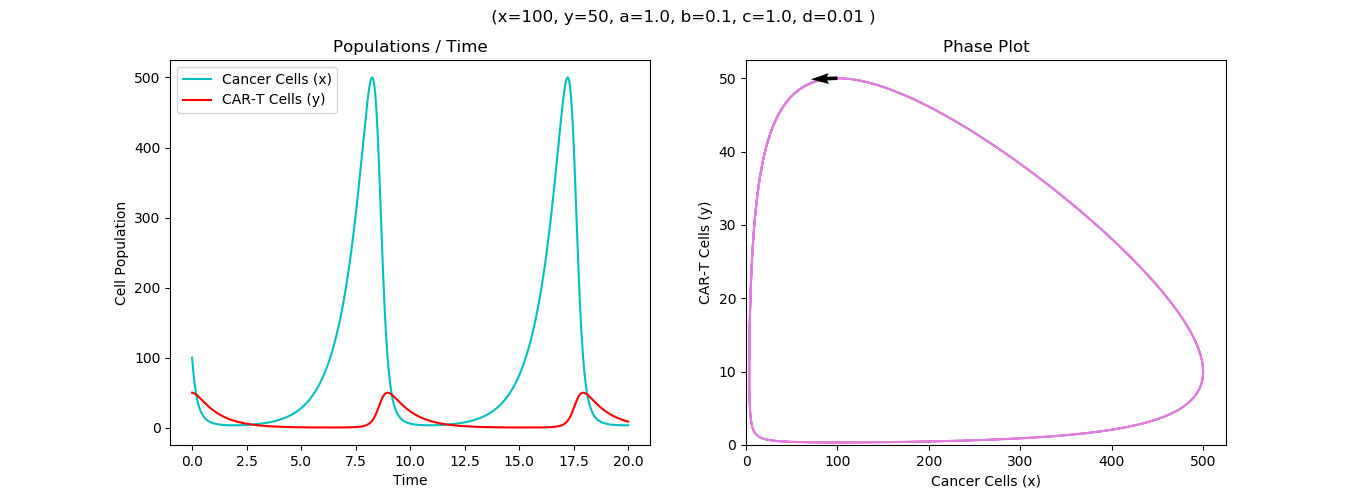
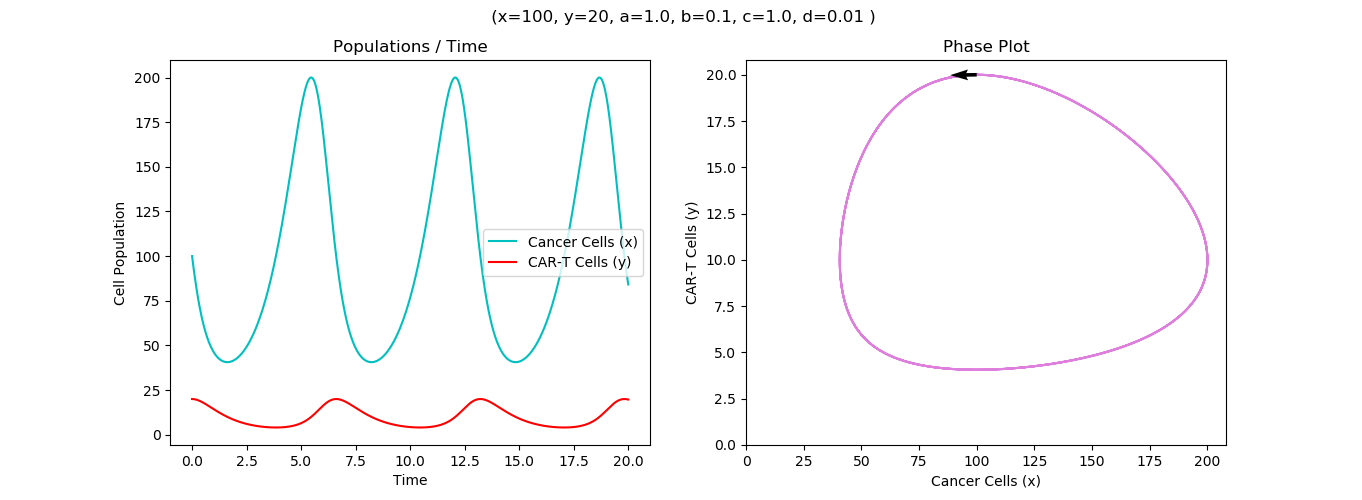

Model Runs on hypothetical Effector to Target ratios E:T, 1:2, 1:5 and 1:20 showing relevant discrepancies in cell counts and killing rates
Our Model has been well fit into our experimental data of cellular counts derived from our sample of Schistosoma associated bladder cancer. We found that the killing rate is inversely proportional to CAR-T doses which could be explained by less opportunity for each CAR-T cell to kill with less cancer cells per CAR T-cells to kill. For instance, with 1:2 and 1:5 E:T ratios, the models resulted in higher rates than 1:20 E:T ratio but requires more time.

Parameter testing plots deducing optimal killing rates with regard to cancer proliferation rates.
Effect of silencing on improvement of Exhaustion

This Run represents a run that considers a factor of silencing as an exhaustion reducing function
Effect of silencing on parameters of 1:2 E:T model with relative improvement of T-cell exhaustion
The CAR net proliferation rate directly proportional to the initial population of CAR T-cells, while the exhaustion rate is inversely proportional to the initial CAR dose. Cancer cell growth was inversely related to the proliferation rate of CAR-T cells, thus on the same CAR-T dose we might encounter variations of immune responses. Thus, balance between T-cells proliferation and exhaustion may be more relevant than the crude killing rates. The model also considered the peaks of cancer cell progression before a successful immune response has been initiated by T-cells. Which could also appear clinically as pseudoprogressions. As shown in the run above, silencing improved exhaustion which show cellular killing rate enhancement and decrease in cancer cell proliferation in comparison with the same 1:2 previous model with no silencing parameters (An exhaustion reducing function) introduced.

This run provides a generated run that describes IL-18 as an additional predator for cancer cells with uniformed expression from CAR-T cells showing relative improvement of killing rate and suppression to cancer proliferation rate
Discussions and Limitations:
Our model represented a mathematical framework for studying the dynamics of CAR-T cells with represetable parameters for cancer proliferation, killing rates, exhaustion, T-cell proliferation and external factors that could affect these parameters such as relative improvement of exhaustion rates by silencing or improvement of killing rates by addition of cytokines. It suggested a 1:2 effector to target ratio as an optimum ratio for desired killing rates of T-cells with considerations of time factor and also revealed dynamics between parameter governing both cell populations in the system.
Our model depends on an in-vitro model for T-cell action which might not be descriptive enough for complex tumor microenvironment. The model still depends on an assumption of uniform mixing between prey cancer cells and predator T-cells. It is also hard to rely upon the accuracy of electrical experimental procedures to differentiate between real dead cell and just detached cell which could release artifacts into considerations of cell counts. The model also does not consider the relative efficacy of silencing vectors or relative efficacy of cytokine production from the intracellular signalling domain of the fourth generation CAR.
References:
1-Kaas, Q., Ruiz, M. and Lefranc, M.-P.IMGT/3Dstructure-DB and IMGT/StructuralQuery, a database and a tool for immunoglobulin, T cell receptor and MHC structural dataNucl. Acids Res., 32, D208-D210 (2004).
2- SAbPred paper: Dunbar, J. et al (2016). Nucleic Acids Res. 44. W474-W478
3-Yifan Song, Frank DiMaio, Ray Yu-Ruei Wang, David Kim, Chris Miles, TJ Brunette, James Thompson and David Baker. High resolution comparative modeling with RosettaCM. Structure. 2013 Oct 8;21(10):1735-42.
4- Wu J, Zhao W, Zhou B, et al. TSNAdb: A Database for Tumor-specific Neoantigens from Immunogenomics Data Analysis. Genomics Proteomics Bioinformatics. 2018;16(4):276–282.
5-Moutaftsi M, Peters B, Pasquetto V, Tscharke DC, Sidney J, Bui HH, Grey H, Sette A. 2006. A consensus epitope prediction approach identifies the breadth of murine T(CD8+)-cell responses to vaccinia virus. Nat Biotechnol 24:817-819.
6-Schneidman-Duhovny D, Inbar Y, Nussinov R, Wolfson HJ. PatchDock and SymmDock: servers for rigid and symmetric docking. Nucl. Acids. Res. 33: W363-367, 2005.
7-Weitzner BD, Jeliazkov JR, Lyskov S, et al. Modeling and docking of antibody structures with Rosetta. Nat Protoc. 2017;12(2):401–416.
8-Kortemme, T., Kim, D.E., Baker, D. Computational Alanine Scanning of Protein-Protein Interfaces. Sci. STKE 2004.
9-Sumbalova L, Stourac J, Martinek T, Bednar D, Damborsky J. HotSpot Wizard 3.0: web server for automated design of mutations and smart libraries based on sequence input information. Nucleic Acids Res. 2018;46(W1):W356–W362.
10-Berendsen, et al. (1995) Comp. Phys. Comm. 91: 43-56.
11-Mestermann, K. et al. The tyrosine kinase inhibitor dasatinib acts as a pharmacologic on/off switch for CAR T cells. Sci. Transl Med. 11, eaau5907 (2019)
12-Yang Liu, Maximilian Grimm, et al . CB-Dock: a web server for cavity detection-guided protein–ligand blind docking. Acta Pharmacologica Sinica, 2019.
13-A mathematical modeling approach to explore kinetics of Chimeric Antigen Receptor (CAR) T-cell Response in glioma: the CARRGO model Prativa Sahoo, Xin Yang, Daniel Abler, Davide Maestrini, Vikram Adhikarla, David Frankhouser, et al., bioRxiv 786020.